Papers
Avolio, ML, La Pierre, KJ, Houseman, GR, Koerner, SE, Grman, E, Isbell, F, Johnson, DS, Wilcox, KR. 2015. A framework for quantifying the magnitude and variability of community responses to global change drivers. Ecosphere 6: art280.
|
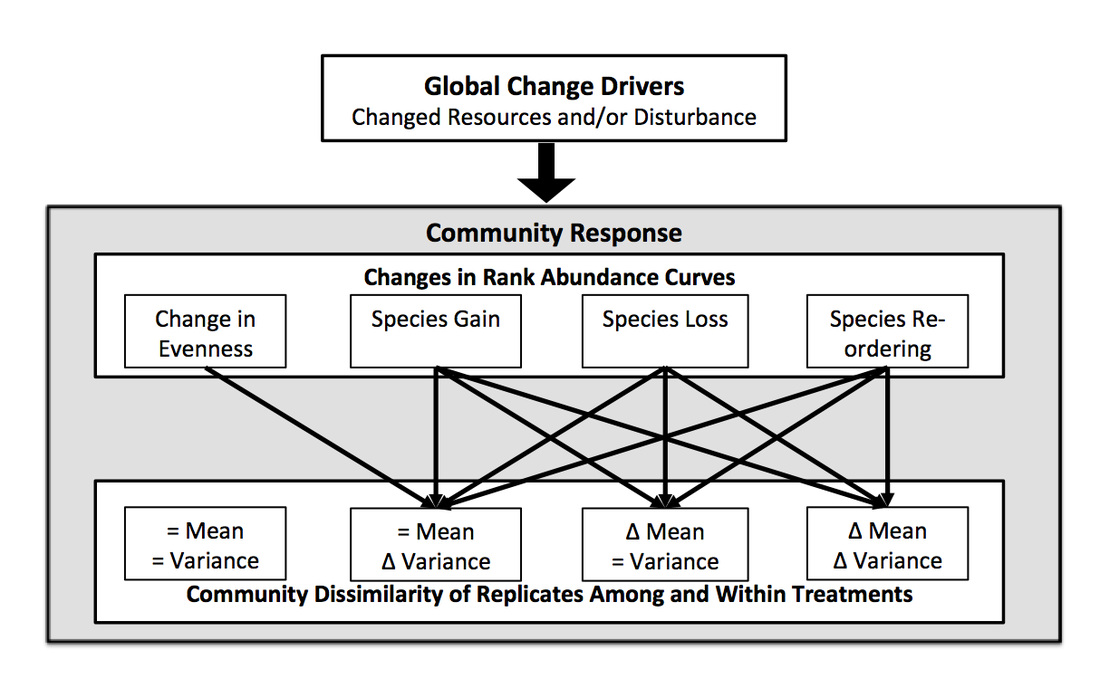
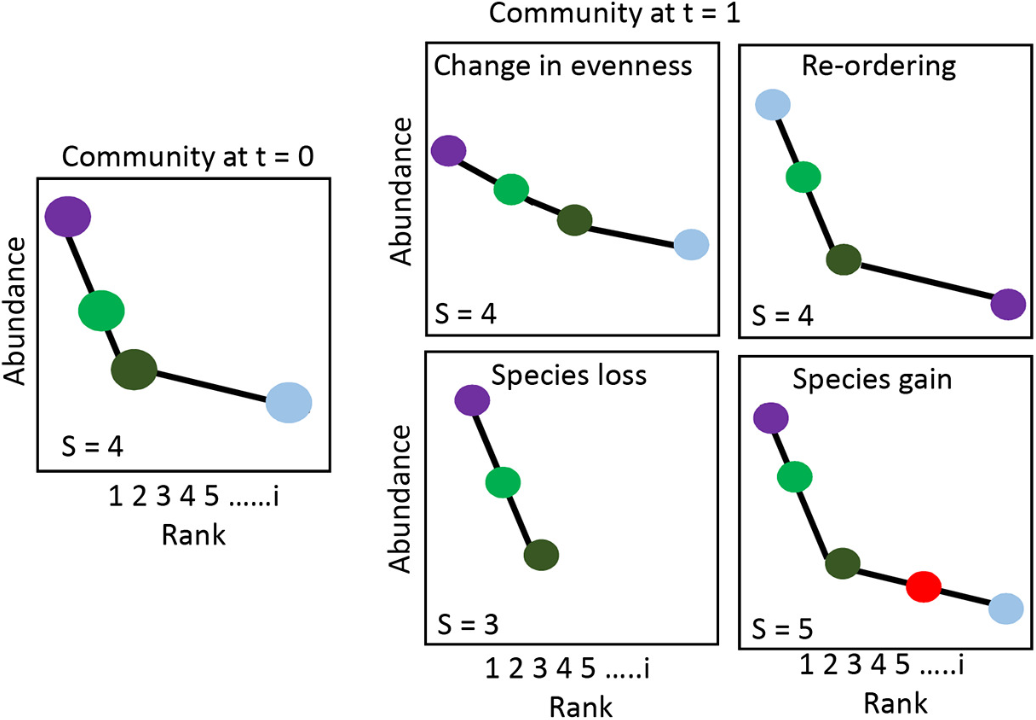
Global change drivers alter abiotic or biotic conditions that influence a wide range of potential mechanisms that change communities. Using rank abundance curves in conjunction with community dissimilarity measures provides insight into the magnitude and predictability of community change.
|
Wilcox, KR, Tredennick, AT, Koerner, SE, Grman, E, Hallett, LM, Avolio, ML, La Pierre, KJ, Houseman, GR, Isbell, F, Johnson, DS, Alatalo, JM, Baldwin, AH, Bork, EW, Boughton, EH, Bowman, WD, Britton, AJ, Cahill JF Jr., Collins, SL, Du, G, Eskelinen, A, Gough, L, Jentsch, A, Kern, C, Klanderud, K, Knapp, AK, Kreyling, J, Luo, Y, McLaren, JR, Megonigal, P, Onipchenko, V, Prevéy, J, Price, JN, Robinson, CH, Sala, OE, Smith MD, Soudzilovskaia, NA, Souza, L, Tilman, D, White, SR, Xu, Z, Yahdjian, L, Yu, Q, Zhang, P, Zhang, Y. 2016. Asynchrony among local communities stabilises ecosystem function of metacommunities. Ecology Letters 20: 1534-1545
|
|
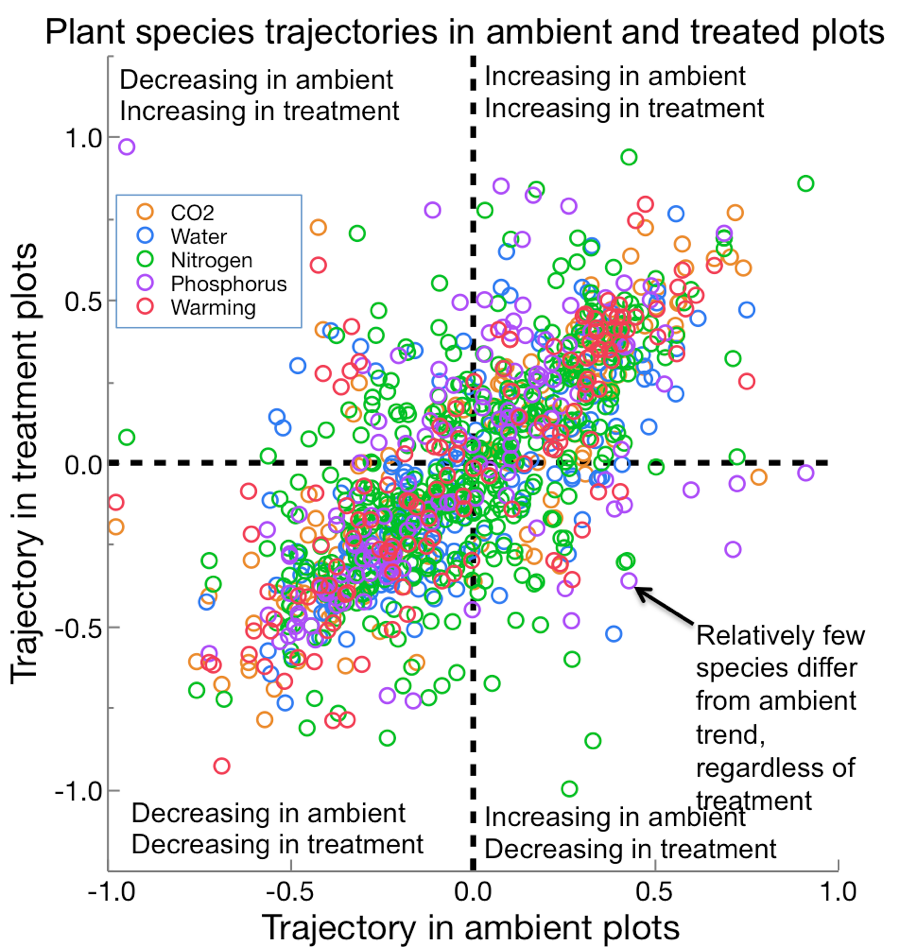
Langley, AJ, Chapman, SK, Avolio, ML, Bowman, WD, Johnson, DS, La Pierre, KJ, Isbell, F, Wilcox, KR, Foster, BL, Hovenden, MJ, Knapp, AK, Koerner, SE, Lortie, C, IN , JP, Newton, P, Reich, PB, Smith, MD, Suttle, B, Tilman D. Ambient changes exceed treatment effects on plant species abundance in global change experiments. Global Change Biology 24: 5668-5679.
|
|
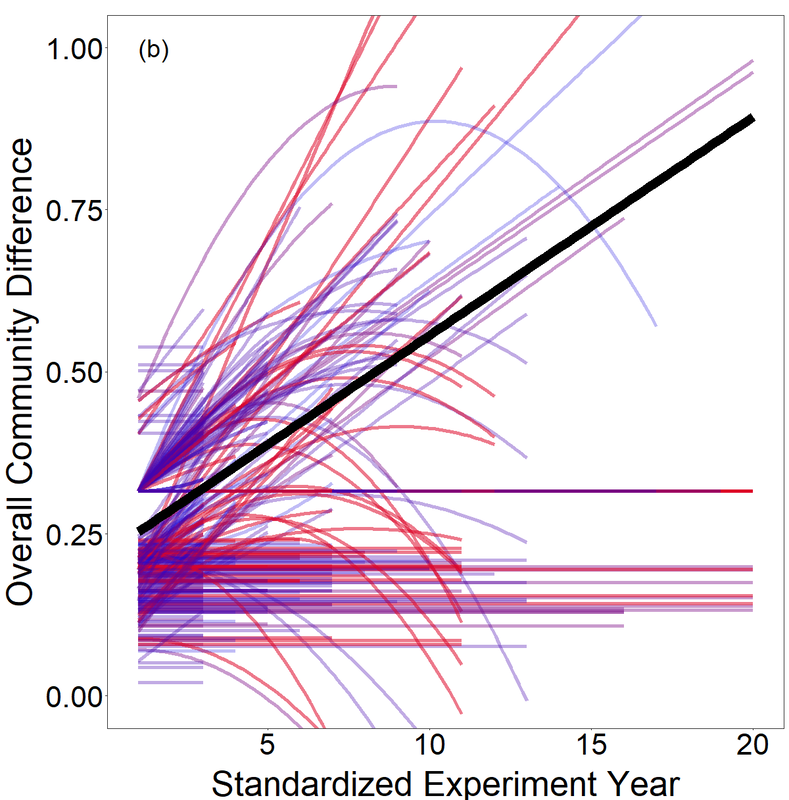
Komatsu, KJ, Avolio, ML, Lemoine, NP, Isbell F, Grman, E, Houseman, GR, Koerner, SE, Johnson, DS, Wilcox, KR, et al. Global change effects on plant communities are magnified by time and the number of global change factors imposed. Proceedings of the National Academy of Sciences USA (PNAS): 116: 17867-17873.
|
|
Avolio, ML, Carroll, IT, Collins, SL, Houseman, GR, Hallett, LM, Isbell, F, Koerner, SE, Komatsu, KJ, Smith, MD, Wilcox, KR. A comprehensive approach to analyzing community dynamics using rank abundance curves. Ecosphere 10: e02881
|
|
Avolio, ML, Wilcox, KR, Komatsu, KJ, Lemoine, N, Bowman, WD, Collins, SL, Knapp, AK, Koerner, SE, Smith MD, et al. Temporal variability in production is not consistently affected by global change drivers across herbaceous-dominated ecosystems. In press in Oecologia
|
|
Wilcox, KR, Komatsu, KJ, Avolio, ML, C2E. 2020. Improving collaborations between empiricists and modelers to advance grassland community dynamics in ecosystem models. New Phytologist 228: 1467-1471.
|
|
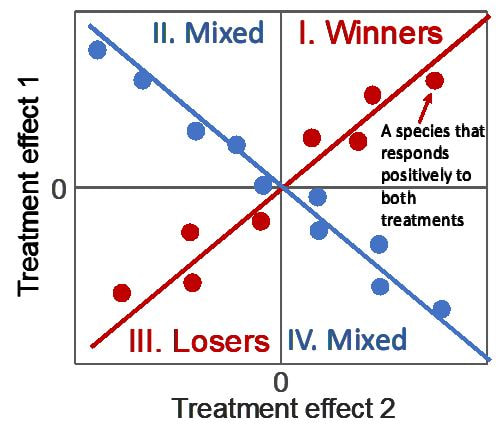
Langley, J, Grman, E, Wilcox, KR, Avolio, ML, Komatsu, KJ, Collins, SL, Koerner, SE, Smith, MD, Baldwin, AH, Bowman, B, Chiariello, N, Eskelinen, A, Harmens, H, Hovenden, M, Klanderud, K, McCulley, RL, Onipchenko, VG, Robinson, CH, Suding, KN. 2021. Do tradeoffs among plant species control response to different global change treatments? Ecology e3626
|
|
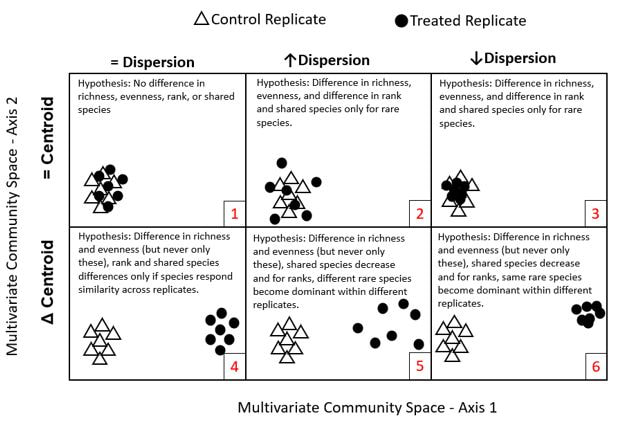
Avolio, ML, Komatsu, KJ, Koerner, SE, Grman, E, Isbell, F, Johnson, DS, et al. Making sense of multivariate community responses in global change experiments. In revision Ecosphere.
|
|
In Preparation
Relating plant community responses to global change drivers to productivity responses
Lead authors: Kimberly Komatsu, Meghan Avolio, Kevin Wilcox, Forest Isbell, Alan Knapp, Sally Koerner, Melinda Smith, Andrew Tredennick
Guidance for improving herbaceous community dynamics in Earth System Models.
Lead authors: Kevin Wilcox, Kimberly Komatsu, Meghan Avolio
Global change drivers alter stability of meta-communities by synchronizing or de-synchronizing plant populations.
Lead authors: Kevin Wilcox, Kimberly Komatsu, Sally Koerner, Meghan Avolio, Yann Hautier, Shaopeng Wang
Grassland plant communities with high rates of background change show greater change with GCDs
Lead authors: Sally Koerner, Kimberly Komatsu, Meghan Avolio
Lead authors: Kimberly Komatsu, Meghan Avolio, Kevin Wilcox, Forest Isbell, Alan Knapp, Sally Koerner, Melinda Smith, Andrew Tredennick
- We have previously found that plant communities respond systematically to global change drivers around the world, however the consequences for functional differences have yet to be investigated in a large-scale synthesis.
- Here we find that productivity responses to experimental global change drivers are initially driven by physiological responses of the existing plant community, followed by reordering of species within a community, and finally species turnover (losses and immigration). This result provides support for the Hierarchical Response Framework (Smith et al 2009).
Guidance for improving herbaceous community dynamics in Earth System Models.
Lead authors: Kevin Wilcox, Kimberly Komatsu, Meghan Avolio
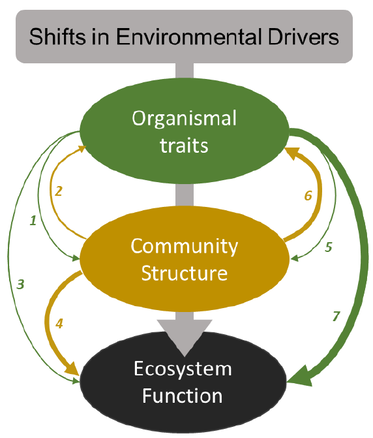
- Earth system models (ESM) are powerful tools to predict ecosystem functioning. However, few ESMs incorporate vegetation change, and those that do, only capture coarse changes.
- Here we demonstrate that current approaches of representing community change into Earth system models often do not match observations, particularly for grasslands, using an example from a long-term resource manipulation experiment in tallgrass prairie. We also discuss commonalities in empiricist and process-based modeler goals and differences in approaches to obtain these goals. Finally, we present a roadmap for better incorporation of occurrences and impacts of community changes into ESMs.
Global change drivers alter stability of meta-communities by synchronizing or de-synchronizing plant populations.
Lead authors: Kevin Wilcox, Kimberly Komatsu, Sally Koerner, Meghan Avolio, Yann Hautier, Shaopeng Wang
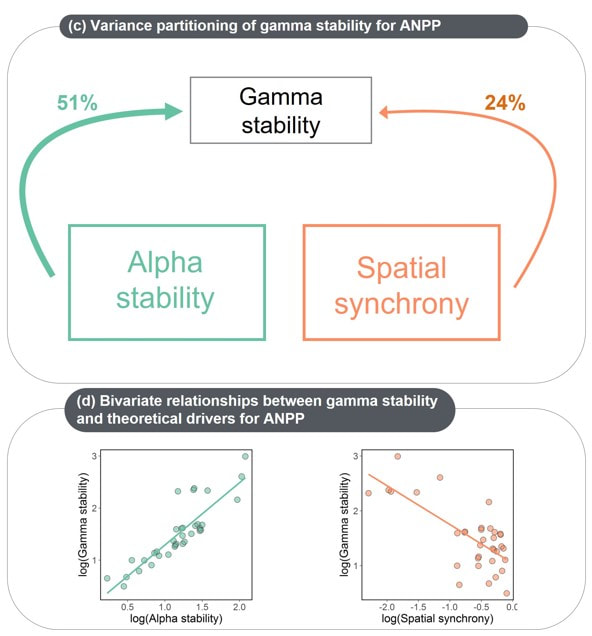
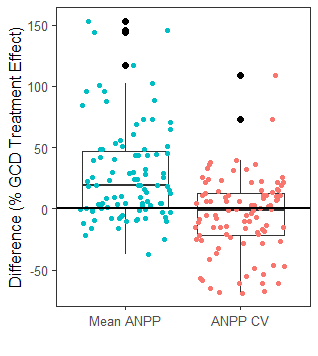
- Ecosystem stability is important for maintaining ecosystem services and are likely to be impacted by global change drivers. We are investigating how global change drivers alter meta-community (beta) diversity, which then scales up impact two important contributors to ecosystem (gamma) stability.
- We find that beta-diversity is a strong driver of population synchrony . This effect is consistent across a wide array of global change drivers.
Grassland plant communities with high rates of background change show greater change with GCDs
Lead authors: Sally Koerner, Kimberly Komatsu, Meghan Avolio
- Plant communities are continuously changing over time. These background rates of change are poorly understood and can have large impacts on system response to global change drivers.
- Some plant communities exhibit higher rates of background change than others, and these communities were more responsive to global change treatment manipulations.
